上位性效应对玉米育种的影响甚微
上位性效应对玉米育种的影响甚微
在遗传学和育种教科书里经常出现上位性效应这个概念,但也经常使育种者感到困惑,甚至使一些研究者产生误解。下面从Hallauer的《玉米育种的数量遗传学》一书摘录部分内容,解释上位性效应在育种中的影响,以飨读者。
人们可以用常见的双列杂交、NCI、NCII、NCIII、部分双列杂交、三重测交、三交、四交等试验设计研究育种群体的遗传参数。这些交配设计在估算方差时都假定无上位性效应。这种假设在各种交配设计中对方差的影响并不明显,与加性和显性效应相比,上位效应的作用比较小,而大多数估算上位性方差的试验是针对广基群体。用自交系杂交形成F2和回交群体的上位性效应似乎比广基群体更重要。因此,通过比较不同世代平均值来检验上位性偏倚就很重要。在p = q = 0.5时,可以估算方差成分和遗传力,但不能把结果推广到其他群体。
对结果进行解释有一定的限制条件,这在各种广基群体是一样的,但自交系杂交形成的群体基因型比广基群体如自由授粉品种、综合种和复合品种的限制更严格。
不管上位性效应有多大,必须表现在基因型的功能上。上位性是两个位点的性状表达,因此,没有理由认为上位性对产量等复杂性状的表达不起作用。尽管上位性效应是肯定的,但需要搞明白上位性效应在总遗传变异中占多大比例。如果上位性方差的比例较小,就可以忽略上位性偏差对选择进度的影响。Buckler等(2009)和Holland(2009)对优系×优系组合衍生的数千份自交系作分子标记,仅检测到不足2%的表型变异属于上位性互作效应(如开花期性状,于是很容易用简单而廉价的混合选择法进行选择)。然而,尽管用近百万植株进行试验,还是没有检测到产量的上位性效应。检测到的上位性效应低得出奇,但在其他生物,确实存在分子互作途径和上位效应。另一方面,Dudley和Johnson(2009)却建议在预测模型中增加上位性,在500个S2家系中,显著提高了预测效果。在总的上位性方差中,只有加性上位效应可以固定,能够被育种者利用。其他大多数分子研究,估算上位性效应没有取得令人鼓舞的进展。然而,从植物育种角度,尽管分子标记数据很少,但上位性很重要。种质选择和准确取样决定了每个试验项目的成功与否。
按数量遗传定义,上位性是纯粹的统计学描述,不意味着生理学功能或表达。上位性方差是位点间(基因间)互作的正交剖分,而一个位点内的三相变化(如:AA、Aa和aa)被剖分为来自回归(

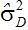
上位性方差剖分为加性×加性,加性×显性和显性×显性三种类型,高层次的上位性包括了其他位点。
尽管有很多方程式估计上位性方差,但总的结果并不令人鼓舞,人们通常无法得到两基因上位性互作成分的估值。例如,Silva和Hallauer发现
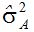
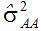
Dudley和Johnson(2009)把上位效应加进偏最小二乘模型,提高了预测籽粒产量和品质的能力。大多数育种群体携带未打破的连锁区段,它们的互作使上位性模型趋于平均,不适用于窄基群体。应该用大样本容量和多点鉴定来提高预测能力。
F检验可用于三交和四交方差分析,检验上位性假设,如果F检验表明变异来源显著,可继续估算遗传方差成分,包括两基因的上位性。例如,在三交分析中有9个亲属协方差,它们有不同的遗传期望。求解均方(或方差成分)的联立线性方程组,然后拟合包含上位性的模型。Wright等(1975)从自由授粉品种Krug黄马齿随机产生60个自交系,通过三交和双列杂交分析估计上位性。通过三交和双列杂交的集合分析,得到9个线性方程,用来拟合包括三基因加性上位性的遗传模型。
不管用哪一种遗传模型和估算方法,
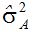
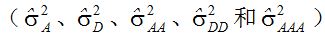
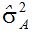
Reid种质在现代玉米育种中的地位非常重要。有人用复杂的交配设计分析Reid黄马齿自由授粉品种,包括五类亲属关系,即:全同胞、半同胞、堂(表)兄(妹)、叔侄和无关系。用七个遗传模型拟合观察值的均方,含三基因的加性上位性。他们用未加权的最小二乘法分析Reid黄马齿品种,未发现上位性效应。用包括
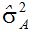

虽然进行了大量研究,包括用大量均方数据拟合含上位效应的遗传模型,都未能获得对上位性方差成分的真实估值。看来,上位性效应在总遗传变异中的分量非常小。
为了获得遗传方差成分估值,需要下面几个假设:(1)随机亲本;(2)二倍体遗传;(3)无连锁效应或达到连锁平衡;(4)无母本效应;(5)环境效应与基因型效应累加。
科学家们用来自群体的随机自交系进行研究。Eberhart等(1966)和Wright等(1971)从原始S0植株选出的自交系样本较小,而Silva和Hallauer(1975)则使用了160个自交系样本,但估计的上位性效应并不比Eberhart等(1966)和Wright等(1971)更成功。把后裔数增加到500个S2,并对伊利诺高油×低油和高蛋白×低蛋白进行测交,尽管概率水平有所提高,但只是略有差别。Wolf等(2000)用B73×Mo17的100个F2植株作父本,按NCIII进行测交,鉴定S1后裔和测交平均值。在B73×Mo17的F2群体中,上位性效应比加性和显性效应小。
连锁效应可能是一个扰乱因素,连锁会增加亲属协方差的上位性系数。在所有研究中,亲本应来自接近平衡的原始群体。玉米中每个染色体上控制一个性状的所有因素似乎不完全连锁。上位性偏倚不是很重要,除非平均重组频率等于小于0.1。因此,在所有变异来源中,连锁效应不应该是一个重要的偏倚来源。上位性成分的系数比加性和显性的系数小很多,这在估计上位性方差成分时是个重要因素。上位性系数小也加大了对标准误的贡献。
比较不同类型杂交种的平均值,在大多数情况下,杂交种是由两个自交系杂交产生的。这些自交系固定了有利的上位性组合。用同一组自交系杂交产生一组平衡的单交、三交和四交种,表明上位效应发生了重组。当用一个单交种产生三交种,用两个单交种杂交产生四交种时会出现重组。对同一组自交系以不同排列方式做杂交组合,进行比较,得到特定自交系组合具有上位性效应的定性证据。这类证据不能定量确定与其他遗传方差比较的相对重要性,但它表明上位性对某些自交系间的特定组合很重要。
上面这些基于数量遗传学分析和分子标记试验结果的讨论表明,产量的基因加性效应占总遗传变异的93%以上,甚至高达97%,还有少量显性效应,而上位性效应非常低,通常不到2%,在多数情况下甚至不到1%。选育自交系的时候,育种者应当重点关注由基因加性效应主导的一般配合力,尤其是在选系的早代,重点测试和筛选一般配合力。到了较高世代,才转向识别和筛选特殊配合力效应。那就和杂交育种衔接起来了。
上述分析还表明,随着种质改良水平的提高,玉米种质的一般配合力呈上升趋势,而特殊配合力效应下降。同时,随着产量水平持续提高,杂种优势持续下降。这是必然趋势。这可能是种质改良的必然结果,同时也为玉米商业育种的技术路线奠定了理论基础。
