RNA干扰育种:硬科学,软工具
基因农业网(Panda)编译:转基因生物(GMO)的合法性在全球范围内还是一个有争议的问题,尤其是在欧洲。不过,表观遗传学领域最新的研究发现,那些没有编码DNA中的遗传信息有可能为基因工程提供更多的选择。今年初发表在《美国国家科学院院刊》(PNAS)上的一篇文章显示,英国某研究小组在没有改动任何DNA序列的条件下,成功地关闭了一个植物基因的表达。虽然该类技术在广泛应用于田间生产之前还有诸多障碍,但各大作物公司已经从中看到了巨大的潜力。(注:表观遗传是指在不改变DNA序列前提下,通过某些机制引起可遗传的基因表达或细胞表现型的变化、某些与经典孟德尔遗传规律不相符的生命现象,常见的机制有DNA甲基化、RNA干扰、组蛋白修饰等。)
RNA干扰及其表观遗传学意义
最初发现DNA时,人们都认为它是携带有机体独特遗传信息的唯一储存介质。然而,上世纪90年代,随着人类基因组测序完成,科学家们发现,基因组中只有极少一部分(1-2%)真正参与组成基因。同时,研究人员还发现了大量的“自私元件”。DNA序列自己拥有在基因组中到处复制以提高生存概率的能力,但“自私元件”的复制活动对有机体来说往往弊大于利:这些寄生性的元件偶尔会插入到有功能的基因序列中去,干扰细胞的正常功能。为了应对这类威胁,宿主细胞进化出了表观遗传学机制来沉默这些自私元件,让它们保持在不活跃的状态。
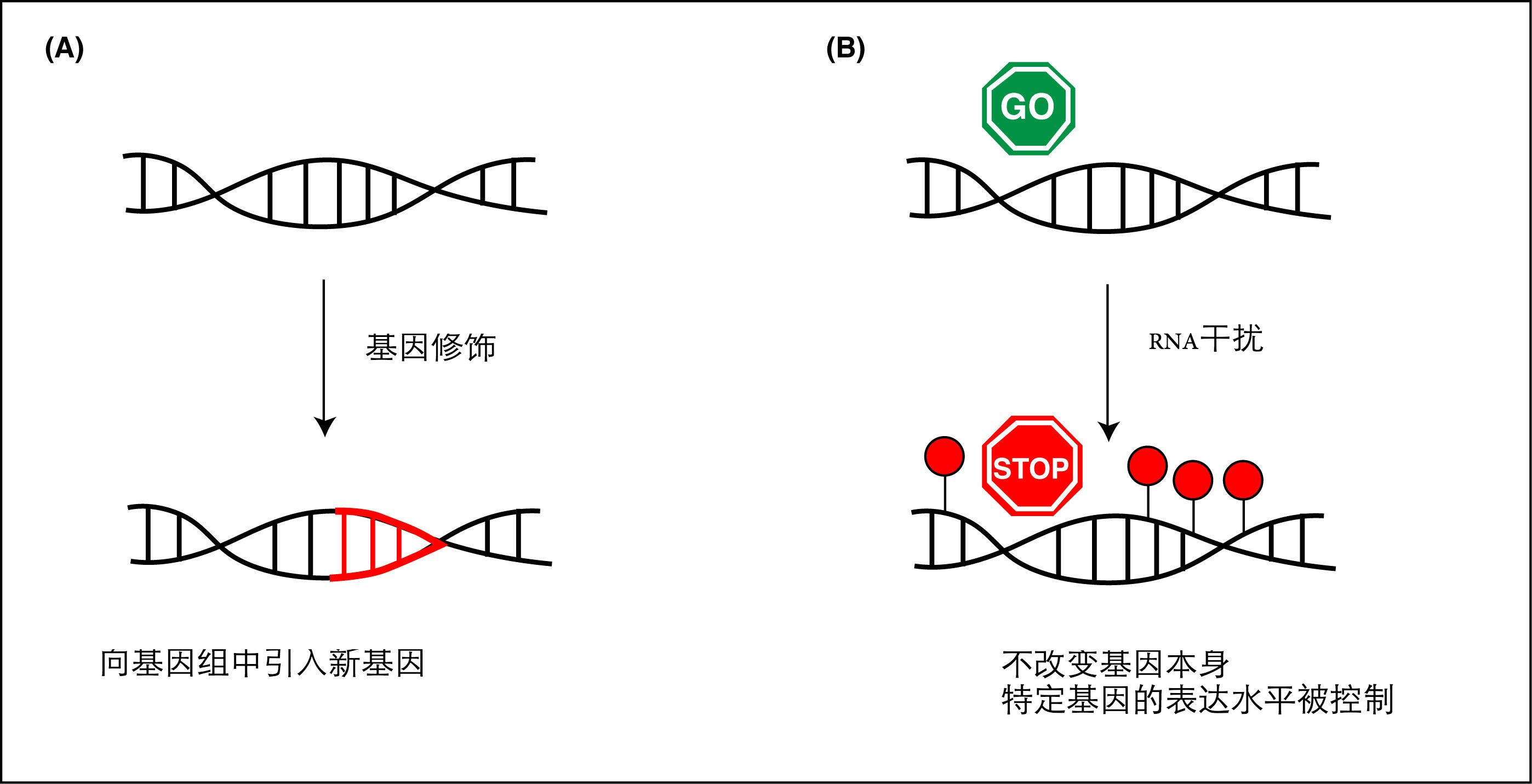
图1. (A)基因工程(改变原有基因序列或是插入一个新的基因)是对某个生物的DNA序列进行改造;(B)表观遗传学修饰不会对DNA序列本身进行改造,而是改变有机体自身对DNA序列的修饰作用。比如RNA干扰,就是一种有机体给自身基因加上沉默标记(图中红色圆点)的机制,从而阻止该基因表达。
与那些不自私的基因一样,自私元件也需要蛋白质来帮助完成自身复制。所有基因表达的第一步都是从DNA转录成RNA,后者作为“信使”分子被“翻译”成蛋白质。如果这些信使RNA(mRNA)在被翻译成蛋白质之前就被降解,那么这些自私元件的功能就会被有效地关闭。1999年,David Baulcombe及其同事发现了这一机制,从此打开了如今已是炙手可热的RNA干扰(RNAi)研究领域的大门。他们发现,如果在体外合成一段能够完美结合靶标mRNA的小RNA分子,并把它注射到植物细胞中去,细胞自身保护机制就会使靶标mRNA降解。不过,如果科学家们继续注射这种小RNA分子,这种干扰机制就会浪费大量能量,因为靶标mRNA源源不断地产生,一旦产生又立即被细胞中的保护机制降解掉。因此,如果持续接触这种小RNA,细胞就会激活一种长期的补偿机制,给DNA序列加上沉默标记,不让它再制造更多的mRNA。这种长期沉默机制被称为RNA指导的DNA甲基化(RdDM)。这一系列标记在DNA上组成了“第二层信息”,科学家们给它取了个名字叫做“表观遗传”:它也是一种信息,但并不像传统的基因那样由DNA编码。那么,这种信息能够被遗传吗?
RNAi现象很快就普及到生命学研究的其他领域,科学家们随后发现,DNA甲基化机制不仅对自私元件有效,而且也会对在那些由于人为转基因操作或是病原体入侵而发生变化的基因起作用。这正是1994年DNA甲基化真正被发现的原因。外源转入的基因被植物体识别为异物,引发DNA甲基化机制使其沉默。不过科学家们一直不清楚该机制是否也能对那些非外源转入基因或是非自私基因(也被称为“无邪基因”,与“自私基因”相对),直到上文提到的PNAS研究中,David Baulcombe和Donna Bond发现了,它能,而且沉默效应还可以遗传。
RNAi也能遗传,沉默无邪基因
Baulcombe和Bond针对一个抑制开花的活跃表达基因,设计了一小段RNA,并借助于病毒载体将小RNA注射到植物体内。如果RNA介导的沉默机制发挥作用,这个抑制基因将会被关闭,植物的开花时间则会相应提前。被病毒感染的植株本身开花时间不会提前,其子代可能还会推迟开花,但子代中确实也存在少数提前开花的植株。再下一代受到的影响更大,提前开花的植株更多。看起来基因沉默效应在逐代增强。在另一个试验中,当植物的抗病毒防御机制被抑制后,研究人员发现,其子代提前开花植株的比例从个别植株变成了几乎100%!由于只有亲本受到病毒感染,得到的子代早花植株都不含病毒。发现在亲本中,该开花抑制基因只在花粉中表达沉默,而叶片并不受影响。这就证实了,虽然基因表达沉默一开始发生在亲本植株中,但对亲本植株而言,该基因的沉默已经太晚了,从已经发育成型的叶片中无法检测到,而这种沉默效应却能够通过花粉传给子代,并在保持两代以上。这是第一次,科学家们证实了植物细胞对“无邪基因”也具有表观遗传学沉默机制,并且沉默效应能够遗传。
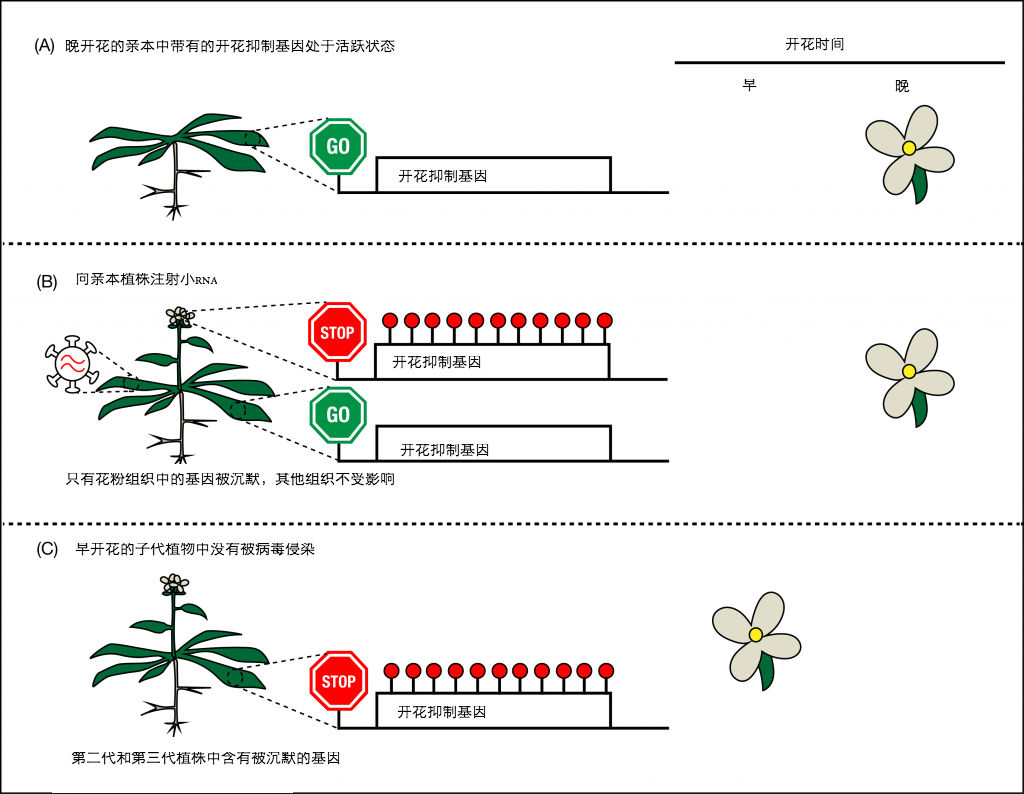
图2. (A)亲本植物中,开花抑制基因处于活跃状态,因此植物开花较晚。(B)感染病毒后,小RNA识别开花抑制基因,并沉默其在亲本花粉中的功能。(C)子代从花粉中遗传了这些被沉默的基因,开花时间提前。
修饰作物性状而非基因组:问题和优势
这一实验室中的重大发现要走向田间应用还有一大堆问题需要解决。这种沉默效应能够持续几代?我们能否提高该机制的功效而不触碰植物自身的抗病毒机制?首先,我们是不是必须在大田里喷施转基因病毒?如果还有其他更好的小RNA载体可选择,这种机制有可能完全取代基因工程。确实,Baulcombe研究小组所进行的实验让我们相信,现在我们完全有能力在不对基因组进行修饰的前提下改变植物的性状。Baulcombe小组的研究对照是控制开花时间的基因,这是一种农艺性状,可能会提高作物对当地适应性。其他一些有意义的性状也值得研究,比如抗旱、耐高温、抗涝等。如果沉默一个基因能够引起有价值的作物表现性状,那么就可以利用这种策略来生产性状改良的作物种子,而无需通过转基因的方法。这就意味着,不必依靠引入外源DNA来获得改良性状了,只需要简单地让植物自身的基因关闭。那么,基因沉默是否可遗传的问题就变得十分重要了。这些种子能够把改良性状遗传到下一代、再下一代吗?会不会某天又恢复到原先的天然状态呢?或者,它们能否像经典的转基因作物那样将改良性状无限期地保持下去呢?对各大作物公司和农民来说,改良性状能够在多个世代中稳定遗传非常重要,决定着这项技术是否具有实际的经济效益。
RNAi除草剂:杂草控制领域的理想黄金国?
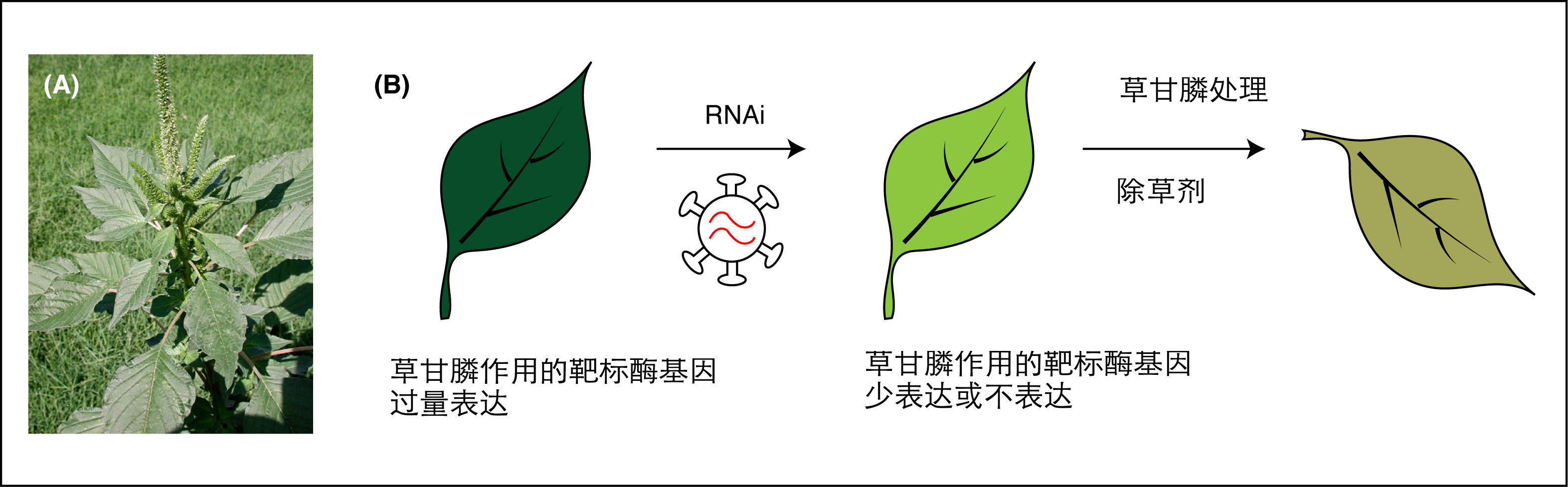
图3. (A)野生的野苋草(Palmer amaranth);(B)由于草甘膦作用靶标酶基因过量表达,使得植物对草甘膦产生抗性;而借助于RNAi技术让该基因沉默,可以让植物恢复对除草剂的敏感度。
RNAi只能让基因沉默,不能增加或增强其功能,因此,DNA甲基化策略就不适用于那些需要从沉默状态激活以发挥功能的基因。由于小RNA并没有向植物体引物任何外源DNA,它们只能作用于植物基因组中已经存在的那些基因。比如,这种策略可能就没办法引入新的除草剂或杀虫剂抗性。不过,沉默现有的抗性基因却有可能实现。我们知道,一旦某类除草剂产品被广泛使用,无需太长时间就会出现对其具有抗性的杂草。比如野苋草(Palmer amaranth)就是一种著名的超级杂草,它对孟山都公司的农达除草剂中有效成分草甘膦具有抗性。抗性的原理很简单,这些杂草大量表达草甘膦作用的靶标酶类,对抗草甘膦的抑制作用,让草甘膦无法发挥功效。如果使用RNAi技术来关闭这类基因的表达,就可以恢复杂草对草甘膦的敏感性。就这样,像孟山都这样的公司开始对RNAi技术感兴趣。
Doug Sammons是在孟山都公司研究草甘膦抗性的高级研究员,在某加拿大农业杂志中声称:“(对杂草控制而言)在过去20年中一度缺少新的探索方向……这确实是一个全新的机遇。”孟山都新的BioDirect技术正是利用了RNAi机制,目前尚在研发阶段,却已经为人所熟知并引发了大量讨论。虽然小RNA的靶标序列能够被精心设计成特异性针对某个特定基因或是特定杂草,但对像欧洲食品安全局或美国环境保护署这类监管机构来说,非预期效应的风险总是一个令人担忧的问题。如果这些小RNA在消化系统中留存下来并进入人体怎么办?如果小RNA在其他基因或其他植物中找到靶标序列了又怎么办?以草甘膦抗性杂草为例,草甘膦作用的靶标酶属于一个蛋白家族,其编码基因序列非常相似,很容易混淆,被小RNA误识别成靶标。这种“脱靶”效应必须被严格控制,它可能会对作物有害。此外,如果“脱靶”效应在子代中保留,并在以后的世代中还能观察到,DNA甲基化的可遗传性就会再一次成为最为重要的问题。
正如Baulcombe和Bond在文章中提到的那样,RNAi策略对各大作物公司来说具有巨大潜力,不过,将这种新兴的、有可能改变游戏规则的技术从实验室研究变成大田中应用的RNAi除草剂或是表观遗传学修饰作物,还需要更多证据来解除公众的疑虑。
Pierre Baduel是有机体和进化生物学专业研究生三年级的学生,从事植物遗传学的研究。作为法国矿业团成员、国家内阁工程师团体成员,他还致力于科学、公共政策和创新关系相关研究。原文链接:http://sitn.hms.harvard.edu/flash/2015/epigenetics-in-plant-breeding/
