作物基因组学与作物科学革命
1中国作物科学的成就与面临的主要问题
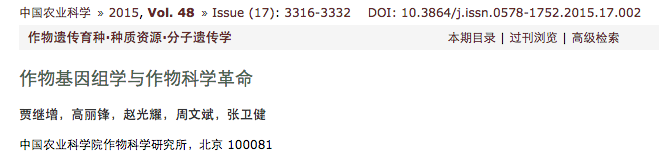
在过去的60多年间,中国主要农作物生产在保障中国的粮食与食品安全、促进国民经济发展与提供就业岗位等方面取得了举世瞩目的巨大成就。中国以占世界9%的土地、6%的水资源养活了占世界22%的人口。近50年来,中国的粮食总产和单产均提高了5倍多(FAO,2014),其中,作物科学的发展对于粮食增产起了关键的作用。迄今,尚未被人们认识的“中国绿色革命”就是一个典型的事例。全世界都认为20世纪60—70年代举世闻名的小麦与水稻“绿色革命”的发源地是国际玉米小麦改良中心(CIMMYT)与国际水稻研究所(IRRI),绿色革命的代表国是印度与巴基斯坦。而实际上中国的小麦与水稻的绿色革命与上述组织(国家)是平行进行的。就矮化育种使用的矮源而言,水稻矮化育种使用的矮源是来自中国的地方品种低脚乌尖与自然突变体矮脚南特,其中,低脚乌尖后来还成了国际水稻所矮化育种的主要矮源;中国小麦矮化育种使用的矮源与CIMMYT不同;就矮化育种时间而言,中国水稻矮化育种始于20世纪五六十年代,还略早于国际水稻所;就增产效果而言,中国粮食产量在1960—1970年10年间年均增长6.8%,等于印度(2.3%)和巴基斯坦(4.4%)2个国家之总和,是同一时期世界上增长速率最高的国家(图1-a,FAO数据)。“中国绿色革命”对于世界粮食增产所作的贡献远高于世界上任何其他国家。因此,如果说CIMMYT与IRRI是绿色革命的发源地,印度、巴基斯坦、菲律宾等国家是绿色革命的代表国,中国则更应该是世界绿色革命的起源地与代表国。中国还是世界上最早培育与推广杂交水稻的国家。该项成果的推广与利用对于绿色革命之后中国粮食产量的大幅度提高产生了重要的作用。长期以来,中国是一个以农业为基础的国家,农业为今天国民经济的高速发展作出了巨大的贡献。虽然目前农业在国民经济中的地位有所下降,但仍然起着举足轻重的作用。中国的种植业是劳动密集型行业,接纳了最多的从业者,在以往的任何时期都提供了最多的就业岗位,保障了中国社会的稳定。
当前,中国的国民经济发展在经历了20余年的高速发展之后,进入了一个稳定持续发展时期。新形势下国民经济的发展对于农作物生产提出了新的要求。
1.1中国粮、棉、油的单位面积产量迫切需要大幅度提高
由于中国是一个人口大国,粮食的需求量巨大。绿色革命、杂种优势利用等成果的推广与利用曾大幅度提高了中国及世界粮食作物的产量。但此后,中国的产量进入一个缓慢增长时期,由原来的年增长近7%降低到1%左右(图1-b),降低了6—7倍之多(FAO数据)。特别需要指出的是,即使这1%左右的增长,也是建立在大量使用水肥资源的基础上,而依靠品种改良取得的进展则更少。例如,黄淮麦区是中国的小麦主产区,也是育种水平最高的地区。对该地区近20多年来育成品种的产量水平进行了统计,结果发现品种改良的遗传进度仅为每年0.7%(图1-c)。由于粮食生产的增速显著放缓,中国近年来进口粮食的速度则成倍增长(图1-d),2013年谷物进口量高达1500万吨,已经由原来的粮食出口国变为进口国。中国目前的粮食自足率早已突破了原定的95%黄线。继续发展下去是十分危险的。与粮食作物的情况相似,中国的油料、棉花生产的现状更不乐观。中国每年植物油消费总量接近3000万吨,其中65%左右为进口;棉花年均需求1015万吨,进口高达350万吨。还应该看到,正是由于进口了大量的油料与棉花,才减少了粮食的进口,否则,中国进口的粮食会更多。由于中国的土地资源有限,加之非农业用地的巨大需求与非农业用地的巨大经济效益,能够保住现有的耕地已是很困难。因此,提高单位面积产量是提高中国作物产量的唯一出路。
1.2中国农作物品种的肥水利用效率迫切需要提高
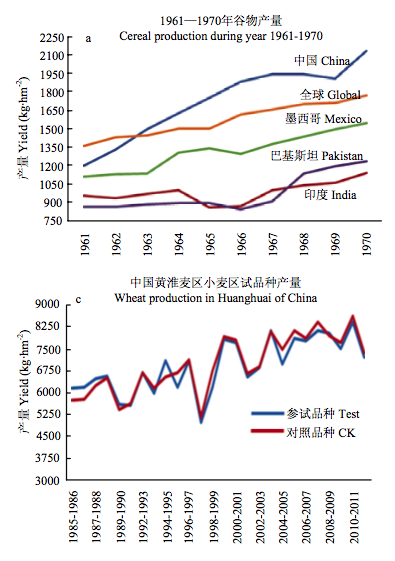

数据来源:a、b 和 d 为 FAO 2014 年统计数据,c 为农业部小麦区试年度资料。
图 1 全球粮食生产状况(a:1961—1970;b:2004—2013)、中国黄淮麦区小麦区试品种产量(c:1985—2011)和中国 谷物进口量(d:2007—2013)
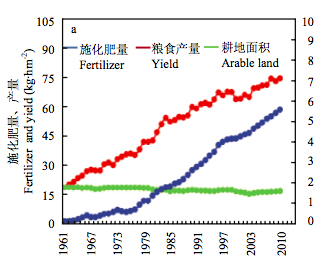
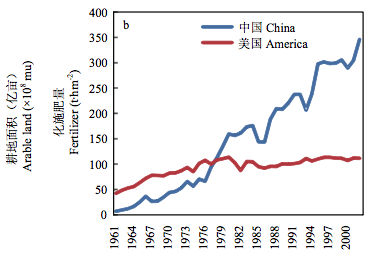

数据来源:a、b 和 c 为 FAO 2014 年统计数据。a:中国粮食产量、化肥用量与耕地面积变化(张福锁提供);b:单位面积化肥用量变化;c:化肥 产量效益变化
图 2 中国和美国化肥施用量和效益比较
中国作物产量的提高很大程度上是建立在肥水大量使用的基础上(图2-a)。当前中国的化肥用量占世界化肥用量的1/3,单位面积化肥的用量是美国的3倍多(图2-b),而产量效益不足美国的1/3(图2-c)。化肥的大量施用不仅增加了投入,而且造成了严重的环境污染。肥料的过量施用造成中国的主要河流、湖泊富营养化,已是不争的事实。中国是一个水资源较为贫乏的国家,人均水资源仅为世界平均水平的1/4。农业用水占全部用水的65%。大量的灌溉用水已经造成河北、河南、山东等地的地下水过度开采,水位下降数十米甚至上百米。不仅严重影响了环境,而且已经影响了人民生活与国民经济发展。因此,有些地方已经不得不限制小麦等农作物的种植面积。上述问题不解决,目前的1%左右的粮食增长水平都难以维持,更不要说进一步提高产量。显然,解决上述问题的唯一出路是培育水肥高效利用的品种以及研究推广肥水高效利用的栽培措施。
1.3中国作物的机械化水平急待进一步提高
中国近年来的农业机械化水平有了较大提高,2014年农作物耕种收综合机械化水平超过61%,较10年前提高了27%,实现了从人力畜力为主到机械为主的转变(2014年11约6日新华网),但与发达国家相比尚存在较大的差距。早在20世纪60年代后期,美、英、法等发达国家的主要粮食作物就实现了从土地耕翻、整地、播种、田间管理、收获、干燥等全过程机械化;20世纪70年代初完成了棉花、甜菜等经济作物从种到收全程机械化。中国与发达国家农业生产率的差异很大程度是由于机械化程度的差距引起的。近年来,由于农村劳动力大量向城镇转移,农业劳动力已严重不足。解决这一问题的唯一出路是实现机械化。同时,实现机械化也是提高中国农业生产率与农产品国际竞争力的主要途径。提高作物的机械化水平一方面靠改良农业机械,另一方面则要靠改良作物品种与耕作栽培制度措施,使之适合于机械化作业,例如培育成熟时种子脱水快的玉米品种、抗倒伏的小麦品种以及适合于机械化收获的棉花、蔬菜与果树品种。
1.4中国农产品的质量与安全急需进一步提高
提高农产品的质量、保障农产品的质量安全是目前人们十分关心的一个问题。一方面,随着人们生活水平的提高,对食品的加工品质与营养品质有了更高的要求;另一方面,由于一些作物病虫害难以控制,造成了农产品达不到安全标准,甚至有毒。2012年,中国长江中下游麦区及部分黄淮麦区赤霉病大发生,不仅造成小麦大幅度减产,而且收获的小麦由于含有超标毒素(呕吐素)而成为人畜都不能食用的“毒麦”。解决上述问题最为经济有效的途径就是培育抗病新品种。
综上可见,按照当前种植业的发展速度已经远远不能满足中国当前国民经济发展的需要,迫切需要开展一场以“更高产、更高效、更优质、更环保”为核心的新的绿色革命。
2大多数作物基因组测序的完成为基于基因组学的作物科学研究奠定基础
以基因组学为代表的各类组学是当今发展最为迅速的一门科学。一门新兴学科的兴起必将促进与之有关的应用学科的飞跃。例如,孟德尔-摩尔根遗传理论促进建立了作物育种学;分子生物学的发展促进了包括分子标记辅助选择和转基因育种在内的分子育种的发展。基因组学将全面促进作物种质资源、遗传育种与作物栽培等学科取得突破性进展,从而促成新的绿色革命。
基因组学研究是近年来世界上发展最为迅速的一个研究领域。随着基因组测序技术的迅速发展,越来越多的主要农作物基因组测序取得了突出进展。继水稻基因组测序完成之后,目前世界各国已完成或接近完成64种作物的基因组测序,其中包括主要粮食作物(小麦、玉米、高粱、谷子等)、经济作物(棉花、大豆等)、园艺作物(主要蔬菜、果树等)等。中国在该研究领域中作出了突出的贡献。在已完成或即将完成的64种作物中,有25种是由中国独立完成或参与完成。基因组测序与分析发现:(1)不同作物虽然基因组大小差异巨大,但其二倍体基因组中的数量相似,均为3万—4万个,基因组大小的差异主要是由于重复序列的比例不同所致;(2)物种的特性与相关基因的数量密切相关。例如高粱的抗旱性与其携带较多的抗旱基因有关,小麦的抗寒性与其携带较多的抗寒基因有关;(3)同一物种不同材料的基因组大小差异可高达20%左右,而人与黑猩猩的差异仅为1%。因此,一个作物物种需要多个参照基因组,特别是杂合基因组。
农作物基因组测序的完成标志着作物科学已经进入基因组学时代。基因组测序不仅第一次在全基因组水平揭示了该物种的组成,而且为之后的种质资源变异组学研究、育种基因组与栽培基因组奠定了基础。
3基因组时代作物科学的重要进展
3.1基因组学对作物种质资源研究的促进——建立种质资源变异组学
育种亲本是作物育种的根本。第一次绿色革命的突破即源于小麦、水稻等矮源的发现与利用;中国杂交稻的突破则是由于“野败”不育系资源的发现与利用。鉴于种质资源的重要性,世界各国及国际组织都十分重视种质资源的收集、保存、研究与利用。目前,世界上保存的各类作物种质资源已达740万份,其中中国保存的种质资源有44.5万份,居世界第二。然而由于研究水平的局限性,这些资源中的重要基因基本没有得到开发,致使育种进展缓慢。基因组学的发展为种质资源研究与开发利用提供了全新的理论、技术与方法,全面促进了种质资源研究的发展。2003年,中国农业科学院在北京组织召开了第一届基于基因组学的种质资源国际学术讨论会(GPGR),目前已召开了3届,标志着国际上对这一新的研究领域的认可。在基因组学的促进下,各主要农作物都先后建立了核心种质。核心种质的建立解决了种质资源数量巨大、难以深入进行研究与利用的难题,并为之后的种质资源重测序、重要新基因的发掘以及资源创新及开发利用奠定了基础。
变异组学(variome)一词首先出现在人类研究中。将其引用到作物种质资源研究中,泛指在全基因组水平研究种质资源变异的种类、分布与功能。变异组学是连接基因组学与育种学的桥梁。变异组学研究的最重要进展就是相继建立了玉米、水稻、高粱、谷子、大豆、黄瓜、西红柿等作物的变异组图谱(即单倍型图谱,hapmap)。变异组图谱的建立为在全基因组水平进行作物的遗传多样性、起源演化、基因发掘以及作物育种学奠定了基础,推动作物种质研究进入一个新的发展时期。遗传多样性是变异组研究的重要内容。各类作物种质资源的多样性研究发现:(1)多样性在物种间的分布极不平衡,异花授粉作物的多样性远高于自花授粉作物;(2)多样性在物种内的分布极不平衡,野生种的多样性远高于栽培种;(3)多样性在基因组内的分布极不平衡,染色体两端的多样性远大于着丝点附近的多样性。起源演化与分类是变异组研究的另一个重要内容,也是此前研究中存在争议较多的一个研究领域。变异组学首次在全基因组水平对资源进行了起源演化与分类研究,为作物的起源演化及分类提供了可靠的分子证据,取得了前所未有的巨大进展。例如,大豆变异组研究将野生大豆驯化的时间提前了2500余年;通过研究水稻基因组变异,Huang等对水稻的起源提出了新的见解。随着研究的深入,可以预计,许多历史上争论了上百年的学术问题将得以解决。特别值得注意的是,变异组学揭示了作物驯化与品种改良中的基因组变异,将为今后有计划地进行品种改良与品种设计奠定基础。
变异组学研究推动了作物种质资源的开发与利用。研究发现,驯化与品种改良不仅对目标基因进行了正向选择,同时由于瓶颈效应也造成多样性的严重丢失,其中包括大量优良基因的丢失,因此,在野生种质资源中存在着大量的、尚未利用的优良变异。重新找回这些千百年前被丢失了的优良基因,是现代作物育种取得突破的重要途径。变异组学正在促进该研究领域的进展。四川农业科学院与四川农业大学利用人工合成小麦为亲本率先培育大面积推广的小麦品种,在区试中较对照增产10%以上,这在多年来的小麦区试中是极为罕见的。英国、澳大利亚等国有望培育出比目前推广品种增产30%以上的“超级”小麦。水稻、小麦、西红柿等作物的研究已经证明,通过培育栽培种特别是主栽品种背景下的野生种导入系,是寻找并利用这些丢失优良基因的有效措施。
在作物基因组测序的促进下,建立起了基于连锁分析、关联分析、比较基因组学、基因表达等一系列的克隆基因的新方法,极大地促进了作物重要农艺性状新基因发掘与功能研究。一批水稻、玉米、小麦等重要粮食作物的产量性状相关基因(穗数、穗粒数、千粒重等)、发育相关基因(抽穗期、光周期、春化等)、抗逆相关基因(抗旱、抗寒、抗盐)、抗病相关基因已经被克隆。作为禾谷类作物模式材料,水稻基因组研究一直处于领先地位,克隆了一大批与产量及产量要素基因,在此基础上,笔者收集了近年来通过图位克隆获得的基因(表3),如调控穗数的基因MOC1、D3、D10、D14、D27、D53、OsSPL14和TAD1,调控籽粒发育的基因LAX1、SPA、Gn1a、OsCKX2、SP1和DEP1等,调控籽粒大小的基因GS3、GS5、GW2、GW5、qSW5和GIF1等,调控抽穗及开花的基因DTH2、DTH3、DTH7、DTH8、Ehd2、Ehd3、Ehd4、Ghd7、Ghd8、Hd1、Hd3a、Hd6、Hd16和Hd17等,调控株型、叶型和根型的基因EP2、IPA1、DWT1、PROG1、Dwarf1、ds-1、OsEBS、NAL1和DRO1等,调控抗病抗逆的基因Xa23、Xa26、Pib、Pi37、SKC1、LTG1、COLD1、qUVR-10和Sub1A等。玉米是继水稻之后完成全基因组测序测定的第二个主要粮食作物,但由于基因组复杂,基因组组装不太好,因而图位克隆进展较慢。2011年以来,已经陆续有7个调控玉米产量、形态建成与抗病的基因通过图位克隆的方法获得。小麦的基因组更大,因而图位克隆的进展也较慢。已经克隆的基因包括驯化基因Q,抗病基因Sr33、Sr35、Lr1、Lr10、Lr21、Lr34、Yr36、Pm3b和Pm6,春化基因Vrn1、Vrn2和Vrn3。小麦全基因组测序近年来有较大的进展,将对小麦基因克隆与功能解析产生重要的促进作用。除主要粮食作物外,园艺基因组学家克隆了园艺和油料作物特殊的基因,如瓜类性别决定基因(CmACS-7)、苦味基因(Bi和Bt)、马铃薯块茎形成相关基因(StCDF1)。由于中国的结构基因组研究基础较好,自然变异与人工突变资源丰富,无论是新基因发掘的数量与质量都走在世界的前列,许多重要的基因都是首先由中国的科学家克隆的(尤其是水稻)。揭示重要农艺性状基因的功能与作用机理、发掘其优异等位基因将为分子育种及其他相关研究奠定基础。
目前变异组研究主要集中在基因组学,尚有一些作物没有开展,或研究不够深入。今后,变异组将会向表型变异组、转录变异组、代谢变异组与蛋白变异组的方向发展。不同层次变异组的结合对作物科学所产生的巨大作用将很快显现。
3.2基因组学对育种学的促进——建立育种基因组学
作物常规育种为作物改良作出了巨大的贡献。迄今为止,生产上的品种基本上都是由常规育种培育的。但是,作物育种的基本理论还是百余年前的孟德尔-摩尔根遗传学理论,上百年来没有大的突破。作物育种在某种程度上还是基于经验的一门艺术。育种的成功往往取决于经验与一定程度的机遇,“效率低、周期长、预见性差”是作物育种长期存在的“老大难”问题。全世界近30余年的实践已经证明,仅靠常规育种难以取得大的突破。基因组学将在育种材料、理论与方法上取得重大进展,从而促进作物育种学这门古老科学取得重大突破,实现由经验到理论的质的飞跃,建立起基于基因组学理论与技术的育种。在基因组学的促进下,有关作物育种的理论取得了如下重要进展:(1)建立了不同作物的重组率图谱,明确了不同作物的重组规律及影响重组率的主要因素,为育种后代的群体设计奠定了理论基础;(2)建立了主要作物的单倍型图谱(hapmap),发现单倍型、重组率与多样性三者存在较密切的关系;(3)在全基因组水平明确了作物驯化与品种改良的主要基因(位点)以及主栽品种与骨干亲本的基因组组成与遗传多样性,为未来的基因组设计育种奠定基础;(4)发现了非孟德尔遗传、表观遗传学及其机理,明确了米丘林遗传学的基因组学基础,从而解决或部分解决了持续上百年的孟德尔与米丘林遗传学派的争论。
作物基因组学正在促进作物育种方法取得重要进展。长期以来,作物育种都是依据表型进行选择,由于表型易受环境条件的影响,因此不仅影响选择的效率,而且影响加代效率。基因组研究发现许多目标性状基因的功能标记或与目标性状基因紧密连锁的分子标记,利用这些标记可以不受环境条件影响,对目标性状进行直接选择,这就是标记辅助选择。近年来,在重测序的基础上,各种作物都开发了高通量的SNP芯片。SNP是继第一代RFLP分子标记与第二代SSR标记之后的第三代分子标记,具有数量多、效率高的特点。SNP芯片既可用于新基因发掘、多样性检测、单倍型图谱绘制,也可直接用于基因组选择育种。目前,水稻、玉米等作物已经研制出了基因组育种SNP芯片。中国已经研制出了有63万个位点的小麦SNP芯片Wheat660,小麦育种芯片正在研制之中。随着越来越多的育种目标基因的发现与育种规律的揭示,未来的基因组育种将对作物改良产生巨大的甚至是革命性的推动作用。
3.3基因组学正在推动作物栽培学取得重大突破—建立栽培基因组学
作物产量、品质与水肥利用效率均是其基因表达的结果。作物育种是通过对基因的选择与重组实现目标性状基因的最佳表达,而作物栽培是通过对环境的调控,来实现作物重要农艺性状基因的最佳表达。二者异曲同工,且作用可相互弥补。作物栽培学家在过去近百年的实践中积累了丰富的经验,建立了比较系统全面的研究体系和技术模式,对作物生产作出了重大贡献。但长期以来,作物栽培研究主要侧重在表型水平,有关栽培措施对基因表达的影响基本不了解,基础理论研究严重不足,已经不能满足现代作物生产的发展需求。栽培基因组学是通过研究环境条件对重要农艺性状基因表达的影响规律,提出合理的栽培措施和实时的生长调控技术体系,实现目标基因的最佳表达。近年来,由于基因组学的飞速发展,在与作物栽培有关的诸方面取得了重要进展,发现了大量的与环境因素有关的QTL,克隆了一批重要的环境相关的基因,如盐胁迫、干旱胁迫、温度胁迫以及营养胁迫如氮胁迫、磷胁迫、钾胁迫等。实际上,大部分基因的表达都是受光、温、肥、水、热、气等环境条件影响。人们在进行多环境QTL分析时,常常发现在不同条件下出现的QTL,而这些QTL很可能就是环境条件影响基因表达的结果。这些基因的克隆将帮助栽培工作者根据基因表达情况来制定最佳栽培措施。反过来,在田间环境下从转录组、蛋白组、代谢组等组学层面上研究不同农艺措施的调控效果,可以更为全面、系统、真实地阐明基因与重要农艺性状的关系,为现代作物分子生物学研究提供新的研究方向。总之,栽培生理学通过与基因组学的融合,必将推进栽培基因组学达到全新的水平。
3.4基因组学促进“小”作物的“飞跃”
据统计,目前人类种植的各类作物共计1500余种,其中,中国栽培的有600余种。就数量而言,水稻、小麦、玉米、大豆、棉花等种植面积很大的“大”作物仅占极少数,而谷子、高粱、食用豆及各类蔬菜等种植面积较小或很小的“小”作物则占绝大多数。这些小作物之所以能够经历数千年的自然选择与人工选择而不被淘汰,说明在这些小作物中还蕴藏着丰富的而且为大作物没有的重要基因,例如,谷子的抗旱性与营养品质突出,其中很可能存在与抗旱及品质相关的基因;木薯、马铃薯与甘薯等薯类作物高光效、水分利用高效与N、P利用高效,其中应该有与上述性状相关的基因。因此,这些小作物也是“特色”作物。然而由于研究力量、投入、历史等原因,小宗作物的研究远远落后于五大作物,制约了这些作物的发展。基因组学大大促进了这些作物研究水平。由于许多小宗作物的基因组很小,方便进行基因组测序与重测序,目前谷子、黄瓜等小宗作物已完成了基因组测序与变异组分析,跨越了大作物花费了近百年走过的表型遗传、细胞遗传、数量遗传历程,直接跨入基因组学时代,赶上了一些大作物的研究水平。这些作物的基因组与变异组研究不仅促进其自身的发展,而且通过比较基因组学与比较变异组学,对于大作物的发展也将产生重要的促进作用。这些作物携带的重要功能基因及其分子机制,对于作物科学的研究也将具有普遍的指导意义。
4目前存在的问题、解决途径与展望
在看到基因组学取得重大研究进展的同时,也应看到当前基因组学研究存在的重要问题。基因组学研究已进行了20余年,发现了上千个农艺性状的QTL,克隆了数以百计的农艺性状基因,发表了许多高水平的论文。然而这些结果却很少在育种上发挥作用。迄今为止,生产上的大面积主栽品种仍然是常规育种培育的。因此,很多人怀疑基因组学是否真的具有应用价值。造成上述现象的原因主要有以下几方面:(1)基因组学研究队伍与育种学研究队伍没有很好地结合,基因组学的研究成果没有有效地转化为育种资源或育种技术;(2)基因组学研究材料与育种学研究材料没有很好地结合,基因组学研究材料难以被育种家所接受;(3)所发现的QTL或克隆的基因大多是在育种上已经被利用,缺乏新的育种价值;(4)许多所谓的“高产基因”实际上是产量构成因素相关基因,并非真正的产量基因。而且这些基因不是在群体条件下,而是在个体条件下获得的,产量构成因素间的负相关没有暴露出来。一旦到了真正的生产条件下,由于相互间的显著负相关而失去应用价值;(5)研究分散、重复,无序竞争,缺乏顶层设计;(6)从理论到应用需要一定的时间。可喜的是已有越来越多的基因组工作者认识到上述问题,越来越多的常规育种家认识到基因组育种是未来育种的发展方向。基因组学家、育种家、资源学家与栽培学家的结合,必将促进作物科学发展的突破与飞跃,促进作物科学进入一个新的历史发展时期。中国应该在这个新的发展时期对作物科学作出中国科学家应有的贡献。
在过去的10年作物科学在基础研究上取得了重要的进展。今后应紧密围绕解决当前中国主要农作物生产存在的“产量增长慢、资源消费高、生产效率低、质量不稳定”等突出问题,综合利用基因组学、代谢组学和表型组学与基因工程等领域的国际前沿科学技术,以实现“更高产、更高效、更优质、更环保”为核心的新的绿色革命为目标,重点开展主要农作物种质资源、遗传育种与作物栽培等方面的基础研究,促进中国作物科学取得重大突破,从而进入一个新的发展时期,促进新的绿色革命率先在中国实现。
致谢:李家洋、朱立煌、杨建昌、陈晓亚、邓秀新、王汉中、李付广等参与了文章的讨论并提出宝贵意见与建议,在此表示感谢。