微生物所揭示细胞形态建成的调控机制

KCBP作为微管与微丝间的连接枢纽,调控表皮毛形态建成。A. 野生型叶片表皮毛形态;B. kcbp-1突变体表皮毛形态;C. KCBP作用模型。
多细胞生命体是由多种多样的细胞组成的,这些形态各异的细胞发挥着各自不同的生理功能。尤其是一些结构与功能均高度特化的细胞的形态建成备受关注,诸如神经系统的基本构成单位神经元细胞,承载着繁衍后代使命、携带精细胞远途跋涉终至雌配子体的花粉管细胞以及千百年来为人类提供天然纤维的棉花纤维细胞等等。细胞骨架(Cytoskeleton)系统,主要为微管(Microtubules, MTs)和微丝(Actin filaments, F-actin),在细胞形态建成中发挥着至关重要的作用;而微管和微丝之间也存在着复杂而有趣的分子对话。但是,目前人们对微管-微丝协同调控细胞形态发生机制的认识还十分肤浅。
植物叶片表皮毛(Trichome)具有减少水分蒸腾、分泌、保护和增强植物抗性的重要作用。模式植物拟南芥叶片表皮毛,是一个很大(约0.5 μm)的、具有3-4个分支的单细胞(见图1A),呈现独特的分支起始(Branching)、伸长(Elongation)、变尖(Sharpening)等生长模式;由于个体较大、位于叶片表面、利于各种研究操作等特征,表皮毛一直是一个研究细胞形态建成机制的理想模型。遗传学研究表明,微管相关蛋白编码基因突变后常常导致表皮毛分支数目的改变;而微丝相关蛋白编码基因突变体分支数目正常、但分支的伸长却有缺陷。有一类zwichel表皮毛突变体呈现非常有趣的表型:表皮毛只有两个分支,其中一个分支伸长和变尖基本正常,但另一分支发生异常、呈现一个短、钝的头;并且表皮毛在叶片上呈有规律的平行列队,短、钝的那一分支总是朝向叶片远轴端(distal end),而尖、细的一头朝向近轴端(proximal end)(见图1B)。上述表型提示ZWICHEL基因应该编码一个同时控制表皮毛分支起始、伸长和变尖的中心枢纽蛋白(Hub Protein),并且很可能同时调控微管和微丝。有趣的是,克隆ZWICHEL基因后发现其编码一个独特的微管马达蛋白——驱动蛋白KCBP(Kinesin-like Calmodulin-Binding Protein);KCBP一直被认为是一个进化中产生的Kinesin和Myosin的嵌合体,因为Kinesin超家族(包括动物界、植物界和真菌界)中唯独KCBP含MyTH4-FERM结构域,而此结构域只在几个动物特有的微丝马达蛋白Myosin中发现过。因此,KCBP是植物中研究最多的一个驱动蛋白,但长期以来关于KCBP调控机制的研究一直没有取得突破。
中国科学院微生物研究所植物基因组学国家重点实验室的孔照胜课题组运用活细胞显微成像技术(Live cell imaging),以三维重构的方式展示了拟南芥叶表皮毛细胞全发育期微管和微丝阵列的时空动态组织:快速伸长的表皮毛分支会形成横向排列、密度趋向顶端的“周质微管环 (Transverse MT rings)”,但其顶端却存在一个“微管缺失区域(MT-depleted zone)”;而恰恰在顶端区域存在一个由周质微丝组成的“微丝帽子(F-actin cap)”结构,帽状结构下面有粗的胞质微丝束贯通基部到顶端(见图1C模式图及动图1、2)。非常有趣的是,KCBP马达蛋白在表皮毛中也呈梯度,且定位于周质微管上,但在顶端特异地富集在“微管缺失区域”(见动图1),因此推测KCBP的顶端富集可能与微丝帽子的组装与维持有关。单分子成像(Single molecule imaging)进而证实,KCBP 独有的N-端MyTH4结构域可结合微管,FERM结构域则可结合微丝。上述研究结果揭示了KCBP作为一个中心枢纽蛋白(Hub protein),协调微管与微丝的时空动态组织,从而调控表皮毛的形态建成。鉴于KCBP在棉纤维中优势表达,该研究对于正在进行的棉花纤维形态建成研究也具有重要的参考价值。
上述研究结果8月19日发表于eLife杂志上(DOI: http://dx.doi.org/10.7554/eLife.09351),助研田娟为第一作者,孔照胜为通讯作者。该研究得到了国家自然科学基金、中科院“百人计划”启动基金及植物基因组学国家重点实验室自主研究课题经费的资助。
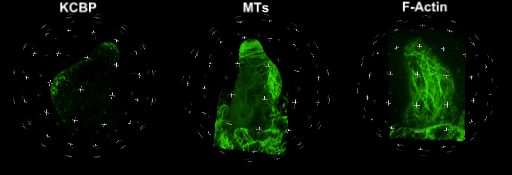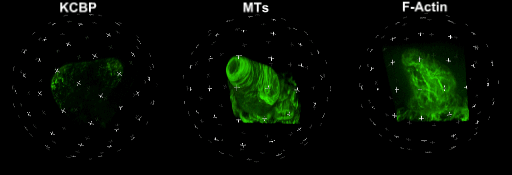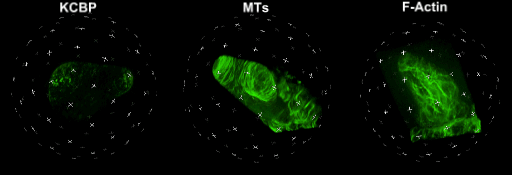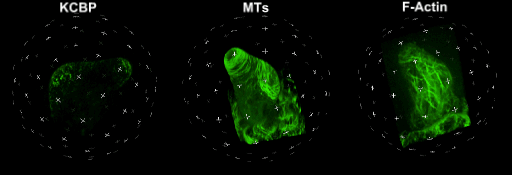
动图1. 处于第一分支起始期的表皮毛中KCBP、微管和微丝的三维空间定位
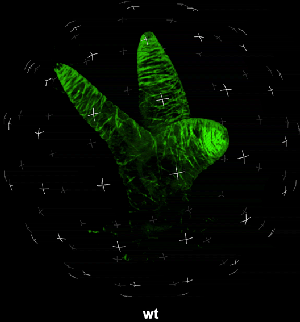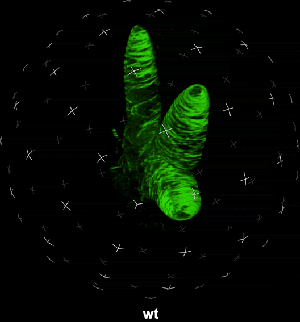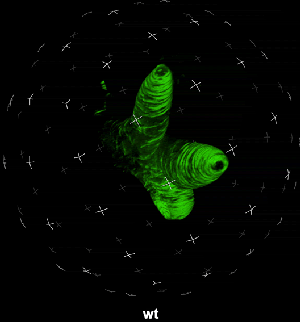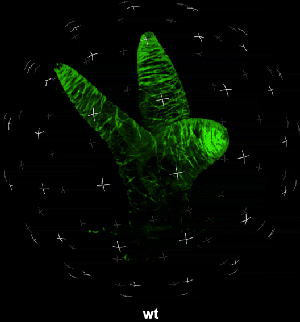
动图2. 表皮毛各个分支快速伸长时微管三维空间分布
(发布:)
