青藏高原所喜马拉雅山南坡阔叶树种林线研究获进展

图1 喜马拉雅山南坡尼泊尔中部9个糙皮桦林线树轮样点

图2 喜马拉雅山南坡糙皮桦高山树线景观

图3 喜马拉雅山南坡糙皮桦树轮横切面显示清晰的年轮界限(未进行染色);其中,最后一个晚材细胞填充有橘黄色的代谢物质,这是区分年轮界限的最重要指示特征。比例尺在A 与B为2 mm, C中为0.2 mm.
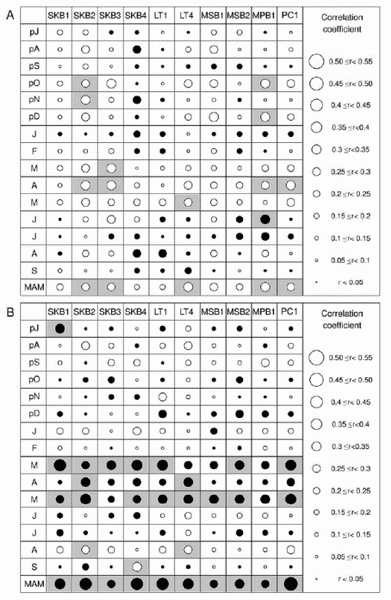
图4 9个糙皮桦林线树轮标准年表以及它们的第一主成分(PC1)与区域月平均气温(A)与月降雨量(B)之间的相关关系。pJ-pD为上年7月至当12月份,MAM为当年3-5月份。实心圆和空心圆分别指示正、负相关系数,阴影区指示相关系数达到p<0.05显著水平。

图5 (A)2004年发生缺失轮个体在不同年龄组中所占的比率,揭示了缺失轮的发生频率与树龄之间不存在显著的关系;(B)所有样点总缺失轮频率的时间序列,显示缺失轮频率呈持续增加的趋势。
国际上普遍认为,高海拔、高纬度林线上树木的生长主要受生长季低温限制,学者们以此为基础提出生长受限假说来解释林线形成机制。中国科学院青藏高原研究所研究员梁尔源及合作者近期研究发现,喜马拉雅山南坡(尼泊尔境内)存在干旱胁迫导致的阔叶树种林线,这对受广泛认可的生长受限假说提出了质疑,为解释高山林线的形成机制提供了新观点。该研究成果已在线发表于美国生态学会的Ecology杂志。
糙皮桦(Betula utilis)在喜马拉雅山南坡形成典型的高山林线(海拔3950至4150米),且多分布在雨影区,海拔2000-3000 米以上降水量随海拔的升高呈下降趋势。该林线最热月(7月份)平均气温9.8°C,5-9月份的平均温度8.2°C,处于全球高山林线分布的阈值温度范围内。沿喜马拉雅山南坡(尼泊尔境内)三个高海拔的自然保护区(郎唐、马纳苏鲁和珠峰),研究人员建立了由9个糙皮桦林线树轮样点组成的网络(图1, 2)。大多数树轮研究基于针叶树种,阔叶树种由于树轮界限不清晰,很少用于树轮研究中。然而,他们发现糙皮桦最后一个晚材细胞填充有橘黄色的代谢物质,为区分树轮界限和交叉定年提供了重要的依据(图3)。对所有9个林线样点的数据分析显示,糙皮桦的生长与3-5月份(季风前)降水量之间存在显著正相关关系;3-5月份的高温通过增加土壤水分的蒸发而加剧水分胁迫,从而与糙皮桦的生长之间呈负相关关系(图4)。同样,西藏纳木错香柏高山灌木(海拔4740-4780米)的生长也主要受生长季早期降水限制(Liang et al. 2012,Annals of Botany, 109: 721–728)。
研究人员发现季风前干旱会造成林线糙皮桦缺失轮的发生,尤其近几十年来的气候异常变暖加剧了林线桦树生长的水分胁迫,造成了缺失轮频率的持续增加(图5)。例如,2003-2004年发生在全球尺度上的极端高温和干旱,造成了林线糙皮桦生长的显著下降,在所采集的树轮样本中2003与2004年分别有17%和18%的缺失轮;发生于1999-2000年的世纪末春季大旱,造成了尼泊尔的农业大减产,林线糙皮桦的缺轮频率(1999为15%,2000为12%)也很好地指示了此极端干旱事件。以缺失轮频率作为指标,2003-2004年是喜马拉雅山中部过去200多年来最严重的干旱事件(图5)。从更长的时间尺度上,糙皮桦树轮序列中连续窄轮的发生,很好地指示了历史上几次主要的干旱灾害事件(例如发生于1756-1768期间的Strange Parallels干旱, 1790-1796的东印度干旱等)。因此,缺失轮频率或连续窄轮记录为糙皮桦林线形成的干旱限制机制进一步提供了有力的证据。
缺失轮的形成是环境胁迫条件超出了树木生长生理阈值的一种适应性响应,高频率或连续缺失轮的发生将会造成树木的死亡。因此,缺失轮频率是敏感的生态指标,可以用来指示气候变化下森林生态系统的脆弱性及其环境限制阈值。
以上研究以中科院青藏高原环境变化与地表过程重点实验室和第三极环境加德满都中心为依托,并得到科技部、基金委和中科院碳专项项目的支持。
标签: 干旱胁迫 南坡阔叶树种 喜马拉雅山
作者:中科院
