陈雷课题组揭示一氧化氮受体sGC的工作机制
发表日期:2019-09-16 09:11PM 阅览次数:
陈雷课题组揭示一氧化氮受体sGC的工作机制
一氧化氮(Nitric Oxide, NO)是一种化学性质活泼的气体,也是一种神奇的信号分子。一氧化氮信号通路在调节血管扩张、神经传递、血小板凝集、免疫、细胞增殖、线粒体呼吸等生理过程中起着至关重要的作用。1998年Robert F. Furchgott, Louis J. Ignarro 和 Ferid Murad 由于阐明一氧化氮是一种信号分子而被授予诺贝尔生理学或医学奖。以心血管系统为例,上皮细胞中的一氧化氮合成酶被上游信号激活后会催化生成一氧化氮。一氧化氮自由扩散至邻近的血管平滑肌细胞中,结合并激活一氧化氮受体——可溶性鸟苷酸环化酶(sGC),进而催化GTP生成第二信使cGMP。cGMP通过激活下游PKG等效应蛋白来发挥生理功能。作为一氧化氮受体的sGC是利奥西呱(Riociguat,用于治疗肺动脉高压)的作用靶点。此外,硝酸甘油,一种从140年前就应用于临床治疗心绞痛的药物,也是通过释放一氧化氮来激活sGC而发挥作用的。
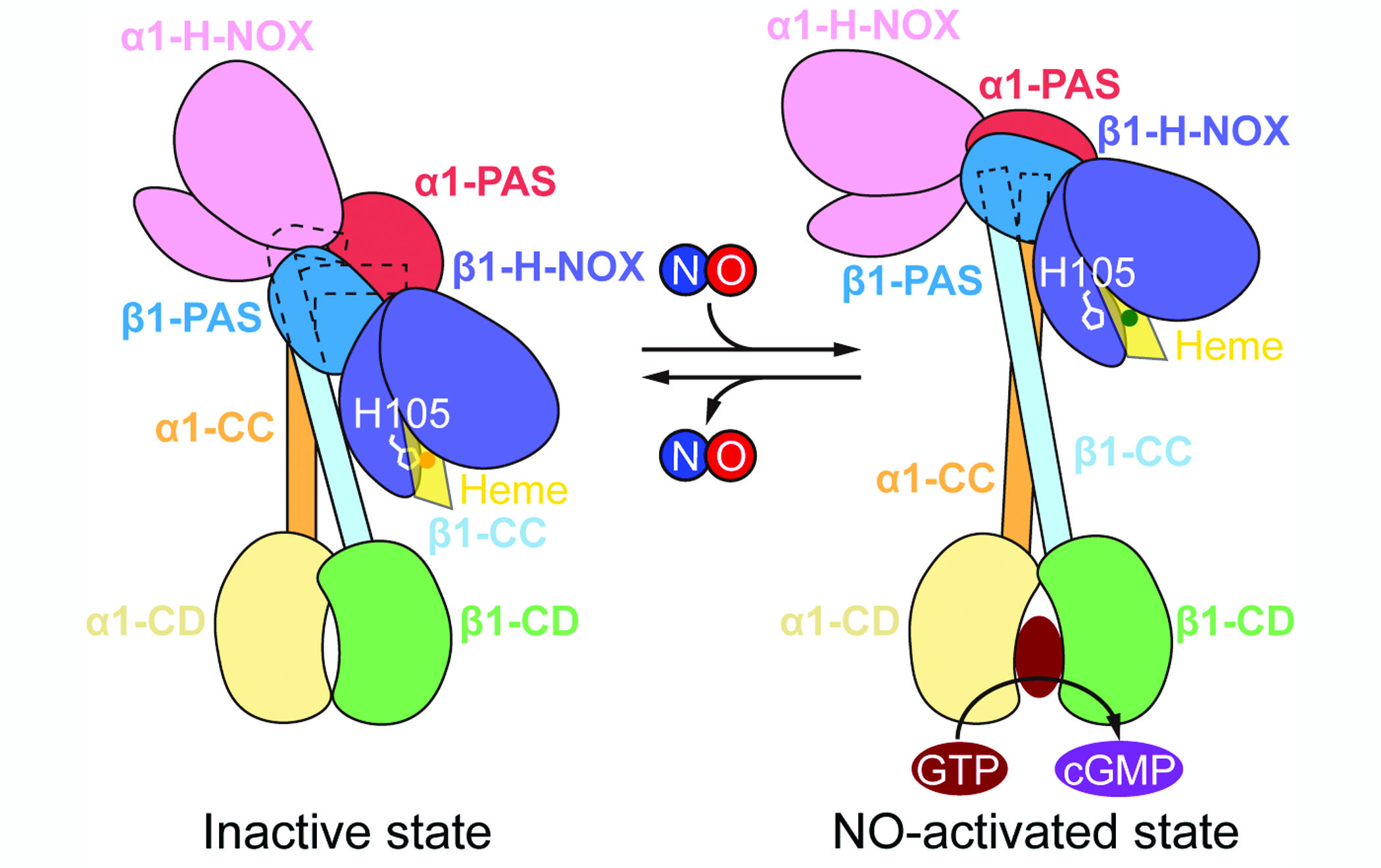
今年是鸟苷酸环化酶发现的50周年。1969年,人们首次检测到鸟苷酸环化酶的活力,发现鸟苷酸环化酶包含两种形式:膜结合的鸟苷酸环化酶和可溶性鸟苷酸环化酶(sGC);在上个世纪80年代,几个研究组分别从组织中纯化出sGC;1988 和1990年,可溶性鸟苷酸环化酶的两个亚基分别被克隆出来。此后的研究证明:sGC是异源二聚体,由一个α亚基和一个β亚基构成。α和β亚基的序列具有同源性,而且结构域排布类似:N端的H-NOX结构域,PAS结构域,CC结构域及催化结构域CD。前期研究表明,β亚基的H-NOX结构域结合了一个亚铁血红素,与NO有高亲和力。该亚铁被氧化成三价铁后酶的活力会降低。虽然此前结构生物学方面的零碎数据对我们理解sGC的机制至关重要,但一氧化氮是如何激活sGC催化活力的仍然是该领域最大的未解之谜。 2019年9月12日,北京大学分子医学研究所,生命科学联合中心陈雷课题组在Nature杂志上发表题目为Structural insights into the mechanism of human soluble guanylate cyclase 的文章,链接见:https://www.nature.com/articles/s41586-019-1584-6。该工作首次解析了人源sGC在无活力状态及一氧化氮激活状态下的高分辨冷冻电镜结构,并结合突变体活力实验,提出了sGC的工作机制。 经过同源蛋白筛选等步骤,陈雷研究组确定以人源α1β1亚基组成的sGC蛋白为研究对象。随后,该研究组制备了四种sGC的冷冻样品:无配体结合状态;血红素铁处于氧化(无活力)状态;一氧化氮激活状态以及H105C组成型激活突变体。陈雷研究组克服了样品制备、数据处理等困难,通过单颗粒冷冻电镜技术获得了上述四种样品分别在4Å,3.9Å,3.8Å及6.8Å分辨率的电子密度,并根据Multibody修正后的电子密度及同源蛋白结构搭建了其原子模型。 无配体结合状态和血红素氧化状态的sGC结构非常相近,被归类于无活力状态。一氧化氮激活状态和H105C组成型激活突变体sGC结构非常相近,被归类于激活状态。在无活力状态下,整个分子出于弯曲状态,其结构可以大体分为:H-NOX和PAS结构域构成的感受模块,CC结构域构成的传导模块及催化结构域构成的催化模块。β1 H105与位于血红素中央的铁离子紧密结合,使β1 H-NOX处于失活构象,而α1 H-NOX的血红素结合口袋被N端螺旋所占据,无法结合血红素。两个亚基的H-NOX结构域在空间上由PAS异源二聚体隔开,并不直接接触。位于整个分子中间的传导模块在靠近N端部分有一个近直角的弯曲,这种弯曲的构象是由β1 H-NOX与PAS及CC结构域复杂的相互作用来稳定的。在无活力状态下,位于C端的催化模块的底物结合口袋处于关闭状态,底物无法结合。二硫键交联实验进一步验证了无活力状态下所观测到的结构域之间的相互作用。 在一氧化氮激活状态下,β1 H-NOX的H105 与Heme中铁离子的相互作用被一氧化氮的结合破坏,导致β1 H-NOX结构域发生构象变化,并且进一步导致β1 H-NOX与周围结构域的相互作用界面发生变化,最终,sGC处于一种与无活力状态差别很大的伸展状态。此构象中,构成传导模块的CC结构域从无活力状态中的直角弯曲螺旋变成了两根完整的伸展的螺旋,该构象变化类似于膝关节的弯曲和伸展。CC结构域两个螺旋之间还发生了70度的相对转动,导致与CC结构域直接相连的催化模块的构象变化:两个亚基的催化结构域之间发生了扭转,底物结合口袋打开,并且在两个催化结构域之间可以观察到底物类似物的结合。通过比较无活力和激活两种状态的构象,我们可以发现β1 H-NOX的构象变化是整个sGC分子结构改变的驱动力,该构象变化通过氨基酸间的相互作用传递至整个感受模块和传导模块,最终到达催化模块。这些结构上的观察与本项工作中报道的突变体活力测定结果相吻合:β1 H-NOX的缺失突变体无法被一氧化氮激活;β1 H-NOX与周围结构域相互作用界面上的突变体的一氧化氮激活大大减弱;传导模块发生构象变化的“关节”位置的脯氨酸突变导致一氧化氮无法激活。 综上所述,本项研究通过强大的结构生物学技术,观测到了一氧化氮激活sGC过程中的构象变化,为深入理解sGC的工作机制奠定了基础。 本项研究主要由北京大学分子医学所博士生康云路和刘锐共同完成,博士后吴惊香参与了部分工作,陈雷研究员为通讯作者。本工作获得科技部重点研发计划、国家自然科学基金委、生命科学联合中心的经费支持。博士后吴惊香先后获得了生命科学联合中心博士后奖学金、北京大学博雅博士后奖学金及中国博士后科学基金的支持。该工作冷冻电镜样品制备、筛选和采集在北京大学冷冻电镜平台上完成,得到了李雪梅、郭振玺、邵博、裴霞和王国鹏等老师的帮助。该项目的数据处理获得了北京大学CLS计算平台及未名超算平台的硬件和技术支持。厦门大学韩家淮教授提供了人源sGC的cDNA,俄勒冈健康与科学大学的David B. Morton教授提供了果蝇和鼠源sGC的 cDNA。 北京大学分子医学所、生命科学联合中心陈雷实验室主要研究膜蛋白特别是与代谢类疾病和心血管疾病相关的重要膜蛋白的结构(http://www.imm.pku.edu.cn/kytd/rcdw/34142.htm)。该实验室此前解析了与糖尿病和心血管疾病相关的KATP通道,与肾病相关的TRPC3/6通道及机械力敏感的atOSCA1通道的冷冻电镜结构。此文报道的sGC工作是该实验室在心血管疾病药物靶点方向的一个分支。陈雷实验室长期招收博士生,希望志于相关方向的研究生加入该实验室。

