我校发表世界首个大规模小麦全基因组重测序研究结果
7月12日,西北农林科技大学旱区作物逆境生物学国家重点实验室在基因组学领域国际知名期刊Genome Biology上发表了题为“Frequent intra- and inter-species introgression shape the landscape of genetic variation in bread wheat”的论文,揭示了六倍体小麦遗传多样性来源于频繁的与野生小麦的种内杂交及与更远缘野草的种间杂交,为了解小麦起源、进化和驯化历史,克服小麦遗传资源同质化、促进小麦遗传改良提供了重要的数据资源。博士生程红、硕士生刘靖、温佳和青年教师聂小军为共同第一作者,姜雨教授、孙其信教授和宋卫宁教授为共同通讯作者。
作为国际小麦基因组测序联盟(IWGSC)成员,西北农林科技大学2016年底率先获得内部共享尚未发表的六倍体小麦参考基因组的信息,开始对小麦遗传多样性来源这一重要命题开展了多学科、多团队联合攻关,对来自全世界的93个野生小麦、六倍体农家种和主栽品种开展全基因组重测序,面对高重复的多倍体基因组分析难题,通过开发新的分析流程,建立了全面的小麦变异组信息,揭示了全球最广泛种植的作物——六倍体小麦各亚基因组的遗传变异,群体内和群体间的遗传分化程度,驯化和改良阶段的受选择信号等全方位的变异系谱和变异来源。
首个大规模全基因组重测序研究提供了目前为止数量最多、覆盖最全的六倍体小麦及其野生祖先的基因组变异集合,特别是筛选出来自多个野生小麦群体和远源物种的大量基因渗入,其中一些渗入片段在群体中出现的频率在驯化或是随后改良过程中发生显著增加。频率改变并与已知数量性状连锁区域重叠,提示这些片段在驯化和改良过程中可能扮演着重要角色。相关结果有助于更深入理解小麦遗传多样性和进化历程,同时为进一步挖掘影响多种小麦表型相对应的功能变异提供了宝贵资源。
研究背景
普通小麦也叫面包小麦(六倍体,AABBDD),大约8000年前在新月沃地由小麦属的栽培二粒小麦(四倍体,AABB)和野生的节节麦(二倍体,DD)自然杂交并加倍形成,由于受到驯化、多倍化以及长期选育的瓶颈效应影响,遗传多样性严重降低。而通过野生种与栽培种之间的杂交,可以有效增加小麦遗传多样性,引入外源优良性状基因。因此,大量野生近缘种在小麦育种的应用具有重要意义。其中,1BS/1RL易位系是应用最广泛、对小麦育种影响最大的标志性案例,黑麦属的黑麦1RL染色体被人工引入普通小麦中,替代小麦1BS染色体,大大提高了小麦的抗病性、品质及产量。“中国小麦远缘杂交之父”李振声院士,通过远缘杂交,将偃麦属的长穗偃麦草的耐旱、抗病等优良基因导入小麦,育成了“小偃”系列品种,其中“小偃6号”是中国小麦育种的重要骨干亲本。但现代育种技术出现之前,农家种中是否广泛存在远缘杂交,却很少有人评估。此外,在小麦起源地新月沃地区域,其四倍体野生祖先种和六倍体普通小麦之间也存在一定频率的天然异交。这些种间和种内的基因交流(渗入)在小麦全基因组上的比例、分布模式至今未得到全面系统评估。
2018年8月,六倍体小麦全基因组的破译为利用全基因组重测序策略研究小麦遗传变异奠定了基础。小麦基因组庞大(约为17 Gb),重复序列含量高(约85%),且普通小麦具有三套相似的亚基因组,组合在一起形成了一个极为复杂的异源六倍体基因组。这些特征都给全基因组重测序、遗传多样性分析和来源评估等研究工作带来巨大困难。本研究对93个小麦及其近缘种材料进行了重测序分析,包括20个野生二粒小麦(wild emmer)、5个粗山羊草(Ae.tauschii)、5个硬粒小麦(durum)、29个六倍体农家种(landrace)和34个栽培种(variety)。另外,还包括24个转录组和90个外显子捕获测序数据的分析。数据分析阶段,研究人员在序列比对、变异检测等过程中进行了长时间的摸索和试错,针对小麦特性专门开发了相关分析流程,最终首次完成了小麦重测序研究工作。

李振声院士与远缘杂交小麦
研究结果
1. 各亚基因组的遗传变异
之前,小麦研究多采用外显子捕获测序或者简化基因组测序等手段,对小麦基因组的遗传变异进行抽样分析,不能全面代表小麦的遗传变异。本研究利用重测序技术,提供了目前为止数量最多、覆盖最全的基因组变异集合,包括84,594,991 个SNPs, 11,628,085 个Indels和205,825个CNVs。全基因组核苷酸多态性(π值)显示AB亚基因组的遗传多样性在从野生小麦到农家种的过程中减少了一半以上(图1b,1c)。D亚基因组的遗传多样性在农家种中非常低,但在栽培种中有所提高(图1d),这可能与和现代育种过程中通过杂交引入更多外源变异有关。此外,基因组变异程度在亚基因组间的分布呈现出不对称性,B亚基因组具有更多的SNPs和CNVs(图1e)。

2. 群体内和群体间的遗传分化
通过群体分析发现,来自西亚黎凡特(Levant,即叙利亚、以色列等地区的统称)南部和北部的野生二粒小麦分成了两个群体,其中黎凡特南部的野生二粒小麦又进一步分成了两支(图2b)。驯化的二粒小麦和普通小麦的AB亚基因组与来自黎凡特北部的野生二粒小麦遗传距离更近(图2c),所以更可能来自黎凡特北部。在形成六倍体普通小麦之后,农家种内部也出现了明显的群体结构。对于AB亚基因组来说,东西方(欧洲和亚洲)的农家种各自聚在一起(图2a, 2d),两个群体之间的分化程度甚至大于六倍体农家种和栽培种之间的分化程度。而中国农家种和世界其他区域农家种的D亚基因组分歧明显,暗示中国农家种的D亚基因组可能存在不同的选育或基因渗入模式(图2f)。
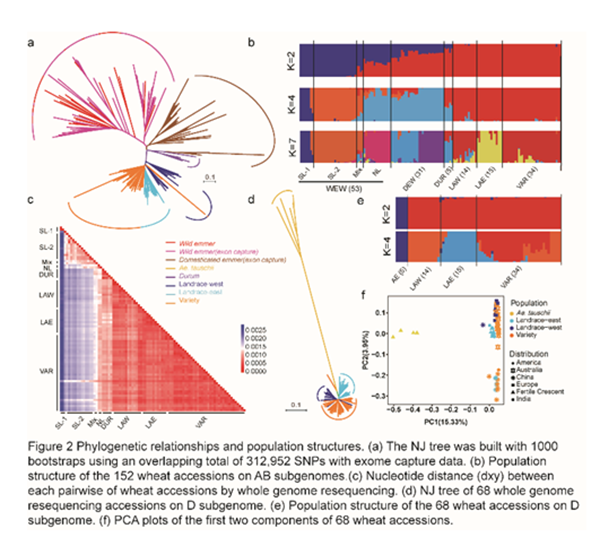
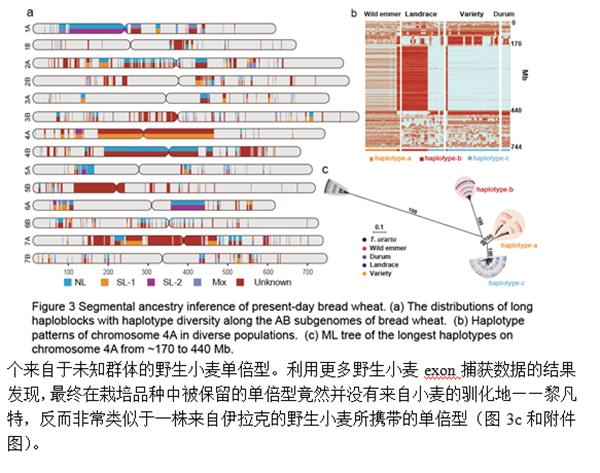
3. 普通小麦同一座位的不同单倍型来源于不同地域的野生小麦群体
为了探究上述不同野生二粒小麦群体对于现代普通小麦遗传多样性的贡献,研究团队进一步系统分析了全基因组上超过1Mb的长单倍型情况。在438个可被鉴定的单倍型区间中,224个区间有Levant北部野生小麦的单倍型(~1164MB),94个区间有来自Levant南部野生小麦的单倍型(~676MB) (图3a)。另外,还有大量单倍型在已有野生小麦样本中无法找到来源,说明可能还有其他野生小麦群体存在。在这些长单倍型中,有一部分处于强连锁的近着丝粒区域,长度达到数百MB (图3b)。特别有意思是,一个位于4A染色体的170到440Mb的长单倍型区间,由三种复等位的单倍型构成,可能来自分歧时间超过10万年以上的三个野生小麦群体。这个座位在农家种中呈现多样性,而现代栽培品种几乎都保留了一个来自于未知群体的野生小麦单倍型。利用更多野生小麦exon捕获数据的结果发现,最终在栽培品种中被保留的单倍型竟然并没有来自小麦的驯化地——黎凡特,反而非常类似于一株来自伊拉克的野生小麦所携带的单倍型(图3c和附件图)。
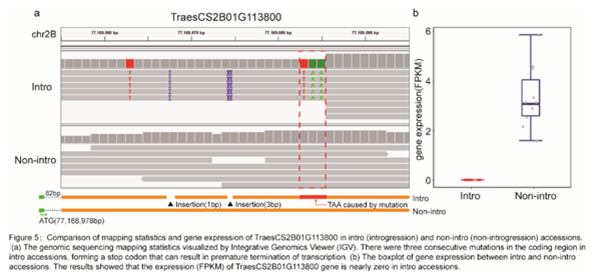
4. 遍布普通小麦全基因组的外源基因渗入
普通小麦基因组除了来自野生小麦的种内渗入,还发现存在远缘物种的渗入片段,在六倍体农家种和栽培种中,分别检测到约709MB和1577MB的渗入片段(图4a)。这些渗入片段极大提高了小麦的遗传多样性,更重要的是引入了优良性状相关的等位基因,大量已报道的产量、抗病、生长发育等相关的QTLs与这些渗入区域重合。研究人员通过基因表达数据,进一步对这些重合区域进行了分析,计算了渗入和非渗入个体差异表达的基因,并从中筛选到一些候选基因、定位了可能的致因变异。如CAM7(钙调蛋白7,TraesCS2B01G113800)基因落在与抽穗期和籽粒重相关的QTLs区域,该基因在拟南芥中是调节光合作者用的转录因子。同时,在渗入个体中发现,该基因通过三个连续的单核苷酸突变形成了一个终止密码子,造成转录提前终止,导致其在渗入个体中不表达(图5)。

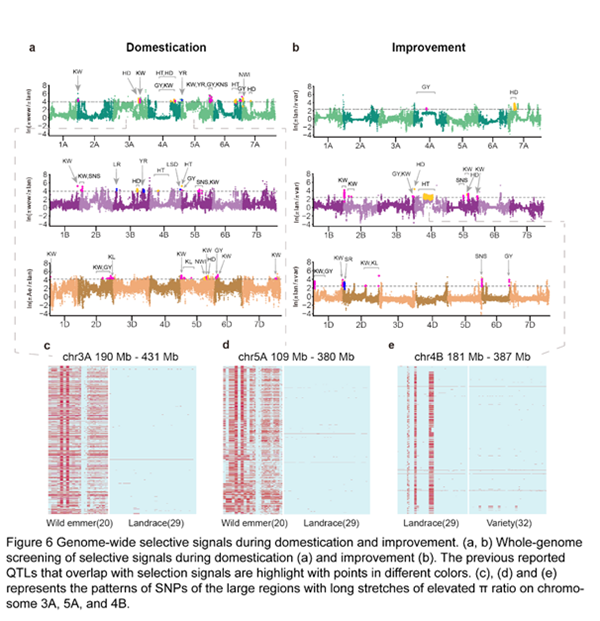
5. 驯化和改良阶段的受选择信号
通过群体核苷酸多态性θπ分析,从野生到农家种的驯化阶段和从农家种到栽培品种的现代育种阶段,研究人员分别鉴定出547 和 438 个选择信号(图6a, 6b),两个阶段的受选择基因分别在昼夜节律和光合作用碳的固定通路富集。值得注意的是,受选择信号在3A, 5A, 4A 和 4B的着丝粒区域延伸超过数十MB。这些区域的单倍型多样性持续降低,29个农家种的3A和5A的近着丝粒区域均固定成同一种单倍型(图6c, 6d),而34个现代栽培品种在染色体4B近着丝粒的200Mb区域也完全固定成了同一种单倍型(图6e),表明小麦在驯化和改良过程均遭受到了遗传瓶颈效应。
该研究得到了西北农林科技大学校基础科研业务费、国家重点研究发展项目(2016YFD0100302,2016YFD0101004)、国家自然科学基金(31771778,31561143005,31822052)和中组部青年国家人才项目的资助。特别感谢西北农林科技大学高性能计算平台和双一流学科群的支持。
原文链接:http://genomebiology.biomedcentral.com/articles/10.1186/s13059-019-1744-x
张晴 终审:闫祖书
